
For this post, I’m using something I published elsewhere. It’s a useful way of looking at the consequences of HIV infection - at least, it has been to me. It contains nothing new; it’s a description of a process that finds a place for many different observations made over the past thirty years. Of course not everyone will attach the same significance as I do to all of these observations.
It’s in two parts, the first is mainly about immune activation and the second about the positive feedback connection it has with HIV replication. This is what I wrote some time ago; I made a few changes for the following two posts.
HIV infection and many other infections caused by a wide variety of microorganisms can have a mutually enhancing relationship that is characteristic of positive feedback systems. It’s useful to explicitly recognize these interactions as positive feedback systems, as this highlights implications for treatment of individuals and for control of the epidemic. It also provides a way of looking at pathogenesis that can suggest further clinical and laboratory studies.
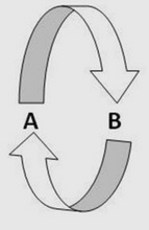
This is an illustration of positive feedback. A stimulates B which in turn stimulates A. In this way the effects of A and B are increased.
Not all co- infections result in a more rapid progression of HIV disease. Many have no effect and a few have even been reported to cause a temporary improvement of HIV disease. This may be the case with measles, scrub typhus and a form of transfusion associated hepatitis. But more often, when an effect of a co-infection has been noted, it has been to promote HIV disease progression.
Worldwide, viruses of the herpes family are probably the most important of the co-infections that interact with HIV in a mutually enhancing fashion. Virtually all adults are infected with some of these viruses that usually exist in a latent or dormant state. They are readily activated in the setting of HIV infection and then can promote further HIV replication by a number of different mechanisms. The previous post describes some interconnections between HIV and the herpes viruses.
In developing nations a range of different endemic infections, depending on geography, may be just as important; many can also accelerate HIV disease progression. Conversely, HIV infection can promote progression of some of these infections.
However there is one characteristic possessed by all HIV potentiating infections. This is their ability to add to the immune activation that is a feature of progressive HIV disease.
By now I think it is generally accepted that chronic immune activation resulting from HIV infection is the major contributor to the pathogenesis of HIV disease. A state of sustained high level immune activation is the basis of the chronic inflammation and immunologic deterioration characteristic of progressive HIV disease.
But what exactly is immune activation?
Immune activation refers to those changes that take place in the immune system when exposed to an infectious agent that allow it to eliminate or control the infection. Essentially, the immune system is activated from a resting state to fight an infection. Generally this process will last for days until the infection is overcome, and usually but not always, is followed by a lifelong immunity to the infectious agent.
However in progressive HIV disease the immune system continues to be activated, sometimes at a high level and it is this sustained immune activation that eventually results in disease. An activated state of the immune system is characterized by differentiation of precursor immune system cells. Differentiation is the process by which these cells develop specialized functions. Examples of cells that have acquired specialized functions are those that produce specific antibodies or those with the ability to target and kill other cells infected with specific microorganisms.
Proliferation of immune system cells is an important characteristic of an activated state. This is usually a short-term response subsiding with control of the infection that stimulated it. But in progressive HIV disease, proliferation is sustained, probably with episodic cycles of further accelerations, and this continued proliferation contributes to the loss of immune system cells.
These cellular changes, differentiation and proliferation, are associated with the secretion of a variety of cytokines. Cytokines are molecules that can change the behavior of cells by binding to specific receptors on their surfaces, for example, causing them to divide. Once released, cytokines not only attach to receptors on other cells but can also come back and attach to the receptors on the cell that produced it. Once attached to their receptors, cytokines set off a cascade of events that result in changes in cell behaviour.
The sequence of events started by cytokine binding will ultimately result in the release of molecules, transcription factors, that attach to specific regions on cell DNA. Some structures present in HIV DNA can also bind cellular transcription factors. . Consequently some cytokines not only change cellular behaviour, they can also change the behaviour of the virus.
Cytokines released in the course of immune activation include those that are associated with inflammatory changes, - the pro-inflammatory cytokines. With respect to positive feedback, pro-inflammatory cytokines including TNF alpha are able to accelerate HIV replication. The positive feedback aspect is this: HIV infection leads to immune activation. Immune activation is associated with production of pro-inflammatory cytokines. These bind to receptors on the cell surface, including HIV infected cells. A consequence of this binding is that a molecule in the cell cytoplasm migrates to the nucleus where it attaches to a specific region of HIV DNA that is integrated into host DNA. This is like turning on a switch that causes many copies of HIV RNA to be made from the integrated HIV DNA eventually resulting in the release of more HIV particles. More cells are then infected and HIV induced immune activation can increase.
A part of the immune system, the innate immune system, responds immediately to infection by recognizing molecular patterns common to different organisms. In contrast, the more familiar adaptive immune system responds to antigens, which are very specific molecular characteristics unique to each organism.
The innate immune system is also activated in untreated HIV infection. Interestingly effects of activation of innate immunity were recognized very early in the epidemic, even before HIV was discovered, and so are among the earliest recognized AIDS related immunological abnormalities. Activated innate immunity is responsible for the large amounts of alpha interferon in the circulation of people with untreated HIV/AIDS, first noted in 1981, the year this disease first came to our attention [1][2] . This is of particular interest to me as I was involved in the discovery of this AIDS related interferon. The sustained presence of such large amounts of interferon had only previously been seen in auto-immune diseases like lupus. At that time the origin of this endogenous interferon was not known. For a period, elevated levels of beta 2- microglobulin were regarded as an adverse prognostic marker. This molecule can be regarded as a surrogate marker for interferon. The association of interferon with abnormalities characteristic of this disease - including low CD4 numbers was also reported in the first 2-3 years of the epidemic [3]. There is an illustration in the last reference showing that high interferon levels are also associated with high levels of a class of antibodies, IgA .
Over twenty years later mechanisms have been discovered that can explain the participation of interferon in the disease process [4].
Interferon appearing in the circulation in untreated HIV disease may even be the first marker of immune activation noted, although not recognized as such when first observed in 1981.
The changes that occur on activation of the immune system are associated with many other markers that can be measured. Different molecules appear on the surface of activated cells. These can be detected and measured, as can the cytokines associated with immune activation.
These measurements can tell us the extent of immune activation. Importantly, the degree of immune activation parallels the rate of HIV disease progression.
Although it is now accepted that the consequences of continued activation and proliferation of immune system cells contribute to the loss of CD4 cells and the development of disease, the precise way it does so is not yet known, although there are a number of different mechanisms that could account for it. The associated inflammation also has adverse effects beyond the immune system, even affecting the cardiovascular system. For more detailed information on these mechanisms here are references to two reviews. [5] [6].
Sustained immune activation is therefore at the heart of HIV/AIDS pathogenesis. It is the sustained nature of the activated state that is critical. Short lived states of immune activation are of course beneficial allowing us to recover from infections. But in progressive HIV disease the process continues at variable rates. Understanding what causes continued immune activation is central to an understanding of the pathogenesis of HIV disease.
What causes Immune activation?
While infection with HIV may start the process, other causes of immune activation almost certainly help to keep it going.
The following all contribute:
1: The immune response to HIV itself. This includes both innate and adaptive immune responses. As noted above, adaptive responses are the familiar specific antibody and cell mediated responses that provide generally lifelong immunity to specific infectious agents. Innate responses depend on recognition of molecular patterns common to several organisms. Some suggest that HIV contributes directly to immune activation through binding of some of its proteins to immune system cells.
2: Microbial products that can penetrate into the intestinal wall as a result of damage caused by HIV. These microbial products then activate immune system cells.
3: Other infections.
Some like active herpes virus infections or the more traditional opportunistic infections can be seen as indirect effects of HIV infection. The word active is underlined because almost all adults will carry latent infections with some of the herpes viruses. Herpes virus infections will precede infection with HIV in most adults.
Other infections also include those that can cause disease in people with intact immune systems like TB or endemic infections in developing nations. Some of these can be more severe in the setting of HIV infection.
Infections that can accelerate HIV replication include those caused by bacteria, viruses, protozoa and helminths.
Those that promote HIV disease progression can usefully be described in three categories.
A: Herpesviruses.
The eight human herpesviruses are described in my last post. Virtually 100% of adults will carry some viruses of the herpesvirus family, usually in a dormant or latent state.
Once infected, individuals carry these viruses for the rest of their lives, usually in an inactive state. All can be periodically reactivated with or without symptoms.
In progressive HIV disease these viruses become active and through a variety of mechanisms, including their contribution to immune activation, promote the replication of HIV. Cytomegalovirus (CMV) may be the most important of the herpesviruses that promote HIV disease progression. It can be part of a positive feedback system in its interactions with HIV.
HIV ? latent CMV infection ?active CMV infection ? HIV
It is not only through their contributions to immune activation that herpes viruses promote HIV replication. In addition to the pro-inflammatory cytokines that have this effect, herpes virus gene products can directly activate HIV if a cell is infected with both viruses. This process, called transactivation works both ways; HIV can also activate herpes viruses.
In addition, herpes infections cause a receptor (Fc) to appear on cell surfaces that allows HIV to enter it. In this way cells that do not possess CD4 molecules can become infected with HIV. Active CMV infections can also exert a mildly immunosuppressive effect.
Herpesviruses, particularly CMV are singled out because they probably play a significant role in the pathogenesis of HIV disease.
The importance of CMV and EBV in contributing to immune activation has been emphasized by Victor Appay in the article cited above in reference [5] dealing with immune activation and inflammation in HIV disease. Here is a quotation from the article: “Hence, sustained antigen mediated immune activation occurs in HIV-1-infected patients, which is due to HIV-1, but also to other viruses (and may be restricted to CMV and EBV)”.
That CMV may play a role in AIDS was suggested by many very early in the epidemic. A multifactorial model for the development of this disease I published in 1983 before HIV was discovered suggested a major role for CMV and EBV [7]. The considerable evidence for a role for herpesviruses, particularly for CMV, did not disappear with the discovery of HIV. The interactions of CMV and other herpes viruses with HIV that have been discovered may now explain their role.
The effects of the anti-herpes drug, acyclovir (Zovirax) on modestly slowing the progression of HIV disease and lowering HIV viral loads were noted in the previous post. These observations have added more evidence that active infection with these viruses can be regarded as part of the disease process for most HIV infected individuals. This is entirely consistent with a model that places HIV and herpesviruses in a positive feedback relationship.
EBV and CMV are very much more resistant to acyclovir than HSV-1 and 2. But it cannot be excluded that this drug did not have some effect in also diminishing reactivations of CMV and EBV. If samples from the trial have been stored appropriately, this can be looked at. EBV reactivation patterns are easily recognized, CMV virus isolation is possible and even detection and quantification of activated T lymphocytes would tell us something.
Valgancyclovir is active against CMV. At the conference on retroviruses and opportunistic infections (CROI) earlier this year Peter Hunt and colleagues reported that this drug reduced CD8 lymphocyte activation in HIV infected individuals. These two bullet points are from their poster:
? Thus, CMV (and possibly other herpesviruses) appears to be a major determinant of CD8+ T cell activation during antiretroviral therapy.
? Given the potential impact of inflammation and immune activation on clinical outcomes (see poster 306), and the potential role of CMV in cardiovascular disease, T cell senescence, and aging, strategies to reduce CMV replication in HIV-infected individuals are worth pursuing in larger trials" .
The poster can be seen by following this LINK.
CMV has an unusual property. Repeated infection with this virus is possible. Just last month a report in Science magazine provided some information on the immunological basis that explains susceptibility to repeated CMV infections. Follow this LINK to read the abstract.
The immune activation that active CMV infections cause can persist even when HIV replication is suppressed by effective antiretroviral treatment. This, taken together with the fact that CMV, when infection is active, can be present in semen means that condom use is also important as a measure to retard progression of HIV disease. .
As mentioned in my previous post, the first publication suggesting condom use as a preventative measure was based on interrupting infection and re-infection with CMV. This is described in the booklet “How to have sex in an epidemic; one approach” written by two of my patients, Michael Callen and Richard Berkowitz.
The relevance of condom use to prevent transmission of CMV is discussed later in the section on implications of looking at HIV disease as one with characteristics of positive feedback systems.
B: Endemic infections:
These are singled out because of their high prevalence in some parts of the developing world.
These infections affect significant proportions of the population, they tend to be chronic and persist in the absence of treatment. The specific infections will depend on geography and many are transmitted by insects. Many of these can also accelerate HIV disease progression, and some also progress more rapidly in the setting of HIV infection [8] [vii].
The term “community viral load” is now familiar. It is likely that in regions where immune activating endemic infections are highly prevalent, community viral loads will be higher than in regions where these infections have a lower prevalence. I don’t know if there have been any studies to look at this interesting question.
C: Other infections:
On an individual level, some episodic infections can promote HIV replication. An acute febrile illness may increase HIV viral loads, but this is a transient effect lasting for the duration of the infection.
Most of the serious opportunistic infections occur late in the course of HIV disease, and may promote even further disease progression.
TB deserves special consideration because of its high prevalence in HIV infection. Susceptibility to TB is increased even at higher CD4 levels. Active TB can then promote further HIV replication thus becoming a partner with HIV in a positive feedback interaction [9] [10] [11].



1 Comment
1 Comment